Днями у журналі Polymer Testing (імпакт-фактор 2,94) вийшла друком нова стаття “The influence of different factors on exploitation properties of nonlinear optical polymeric materials based on an epoxy matrix doped with flavonoids” за авторства групи дослідників, серед яких і викладач нашої кафедри Ольга Неділько. Вітаємо Ольгу Павлівну зуспішним завершенням цього тривалого дослідження!
Продовження фахових розповідей про збудник COVID-19 (а також його родичів та друзів) від автора університетського підручника “Вірусологія”, доцента нашої кафедри Сергія Шамрая (доступний за посиланням). Читайте також першу, другу та третю частини на нашому сайті, якщо Ви досі цього не зробили.
Ось і наступний (і можливо останній) есей про SARS-CoV-2 і хворобу з дивною назвою COVID-19. До цього есея мене спонукало відео на ютубі, яке мені надіслали.
З нього я зокрема з подивом дізнався, що на початку 20-го сторіччя була епідемія ВІРУСНОГО захворювання – черевного тифу, проти якого робили ПРОФІЛАКТИЧНІ ЩЕПЛЕННЯ! Ну, з відео я ще багато чого цікавого дізнався, хоча додивитися до кінця не зміг. Крім цього відео, є багато дописів, автори яких маніпулюють статистичними даними і доводять, що нема насправді смертей від цієї хвороби. Або навіть і сам новий коронавірус є вигадкою, або що вірус передається через 5G, і взагалі все це розробка чи то жидо-масонів, чи то ілюмінатів, чи то Всесвітнього уряду на чолі з Соросом, чи то їх усіх разом. А може, насправді це їхня вигадка, щоб зруйнувати економіку і все забрати собі.
Сперечатися з цим усім – марна праця. Якщо людина у це ВІРИТЬ – як можна переконати, що її віра марна? При цьому, чим більша дурня, у яку вірить людина, тим міцніша її віра. Тому я не збираюся дискутувати щодо окремих пунктів таких дописів.
Перше, що хочу сказати. SARS-CoV-2 дійсно існує, як і інші шість бетакоронавірусів, які інфікують людей. Він виник внаслідок мутацій і рекомбінацій у вірусу кажанів, на які впливав природній добір, це беззаперечно. Дуже гарний пост на цю тему зробив пан Олександр Зіненко.
Друге, чи дійсно від цього вірусу помирає так багато людей? Мені тут важко щось сказати, але якщо вважати, що летальність від нього не вище, ніж від сезонного грипу, то виглядає так, що наприклад у Італії і Іспанії придурювались, що в них бракує місця в моргах. У минулому році теж льодовий палац у Мадриді використовували як тимчасовий морг? Трамп також не вірив у коронавірус, але був вимушений ввести карантинні заходи. У Нью-Йорку і минулого року також померлих ховали у братських могилах? Але це звісно не вірус Ебола і навіть не атипова пневмонія. Тому я б сказав так: не треба панікувати, але не треба і легковажити. І деякі обмежувальні заходи, хоча і набридлі, але є необхідними на деякий час.
Трете, вакцина від SARS-CoV-2. Зрозуміло, що найкраще було б зробити вакцину (і отримати мільйон зелених від нашого, перепрошую, президента). Але досі невідомо, чи формується стійкий набутий імунітет до нього. Є погана новина – у Японії і Південній Кореї фіксували випадки повторного зараження SARS-CoV-2 у людей, які одужали після першого зараження. Але поки що панікувати не слід. У людей завжди трапляються випадки, коли після вакцинації (як і після перенесеної хвороби) не встановлюється достатній рівень набутого імунітету. Інша можливість – у цих людей вірус десь сховався у організмі від імунної система, а потім чомусь реактивувався (але спеціалісти визнають, що не розуміють, де він міг ховатися і як реактивувався). Дійсно погана новина буде, якщо антигени SARS-CoV-2 можуть достатньо швидко мутувати без зниження інфективності і вірулентності (як наприклад у вірусу грипу). І взагалі, що зараз можу сказати я стосовно набутого імунітету, якщо виконавчий директор ВООЗ Майк Раян нещодавно заявив: «Що стосується видужання, а потім (можливого) нового зараження, я гадаю, що ми не маємо відповіді на це запитання». Лякає тільки, що російськи спеціалісти пригрозили влітку розпочати випробування вакцини проти SARS-CoV-2. Враховуючи їхню кваліфікацію, після початку випробування можна очікувати нового спалаху хвороби. Але, можливо, вони не будуть використовувати у вакцині послаблені або вбити вірусні частки. А з білками-антигенами хай експериментують.
Четверте, ліки від COVID-19. Якщо вакцинація, тобто активна імунізація проти SARS-CoV-2 поки що є віддаленою перспективою, то пасивна імунізація, тобто введення плазми крові осіб, які одужали від COVID-19 (так званої реконвалесцентної плазми), видається перспективним. Ця плазма крові містить специфічні до вірусних часток антитіла, і введення 200 мл плазми на тлі підтримуючої терапії не викликало негативних наслідків і покращило стан хворих. Принаймні, так пише Панас (Proceedings of the National Academy of Sciences of the United States of America, PNAS), якому я довіряю. Хоча автори цієї статті усі китайці.
Що стосується противірусних препаратів, згадуються і ліки проти малярії, і препарати-інгібітори протеаз (пам’ятаєте, розрізання протеазами білка S критично важливе для потрапляння вірусної частки в клітину) і багато інших. Але все це на стадії випробування. І навіть якщо були б вже препарати з доведеною ефективністю для лікування COVID-19, у мене є принцип – не давати нікому ніяких рекомендацій щодо лікування вірусних інфекцій. Я не лікар, не давав ніякої клятви Гіппократа і якщо експериментую зі здоров’ям, то із своїм власним. Цю стару мавпу вже нікому не жаль, навіть мені самому.
Згадаю тільки два класи противірусних препаратів, цікавих з точки зору загальної вірусології. Перший клас – це аналоги нуклеозидів, які у інтернеті чомусь часто звуть аналогами нуклеотидів. Але нуклеозид перетвориться на нуклеотид тільки після фосфорилювання. Сенс використання таких препаратів полягає в тому, що аналог нуклеозиду перетворюється на аналог нуклеотиду тільки чи переважно в тих клітинах, де є вірус. Тут він і блокує синтез нуклеїнових кислот, вставляючись у ланцюг нуклеїнової кислоти, що синтезується, замість звичайного нуклеотиду.
Подальшій синтез нуклеїнового ланцюга зупиняється. Деякі з таких препаратів є на диво безпечними і не мають негативних ефектів на здоров’я. Низка аналогів нуклеозидів використовують проти РНК-геномних вірусів, вони блокують синтез РНК РНК-залежною РНК полімеразою. Але цей клятий SARS-CoV-2 певним чином є винятком. Для того, щоб зрозуміти, у чому полягає виняток, зробимо маленькій ліричний відступ. Реплікація ДНК виконується ДНК-залежною ДНК-полімеразою, яка має здатність виправляти невірно спаровані (некомплементарні) основи. Ще кажуть, що ДНК-залежна ДНК полімераза здатна до пруфридінгу (proofreading). А от РНК-залежна РНК полімераза у загальному випадку до цього не здатна, і саме тому віруси з РНК-геномами мутують і еволюціонують набагато швидше, аніж віруси з ДНК-геномами. Але у коронавірусів є клятий білок 3′-5′-екзорібонуклеаза (ExoN), який здійснює виправлення помилок спаровування основ і зокрема видаляє аналоги нуклеотидів. Вірогідно, завдяки саме цьому білку коронавіруси і мають найбільші РНК-геноми серед вірусів. Показано, що саме цей білок SARS-CoV-2 видаляє аналоги нуклеотидів, які утворилися з аналогів нулеозидів, що робить їх неефективними. Тільки один аналог нуклеозидів, ремдесивір (remdesivir), відомий також як GS-5734, виявив активність проти SARS-CoV-2. Цей препарат є аналогом аденозину. Таким чином, потенційно ремдесивір, який спочатку був розроблений зокрема для боротьби з вірусом Ебола, може бути перспективними ліками і для COVID-19. Кому цікаво ознайомитися з статтею на цю тему, ось вона.
Другий клас противірусних препаратів, про який я хочу згадати, це чудернацькі агенти, які діють на молекулярному рівні, і я не зовсім впевнений, що їх реально будуть використовувати, принаймні скоро.
Перший з них – це противірусна система CRISPR бактерій і архей (від Clustered Regularly Interspaced Short Palindromic Repeats). Треба зазначити, що ця система, як зараз стало відомим, є не тільки противірусною, але й приймає участь у регуляції експресії власних генів бактерії. CRISPR-касети виявлені в геномах багатьох бактерій і більшості архей. Повтори мають довжину від 24 до 48 пар нуклеотидів; вони мають бівалентну симетрію, але, як правило, не є істинними паліндромами. Повтори розділені варіабельними ділянками ДНК, спейсерами, приблизно однакової довжини. Спейсери відповідають по нуклеотидній послідовності певним фрагментам ДНК чужорідних генетичних елементів (протоспейсерам). У зв’язку з цим було запропоновано і потім показано, що послідовності, що розділяють повтори, походять від послідовностей геномів бактеріофагів і, відповідно, забезпечують захист клітин від інфекцій. Вище CRISPR-касет (upstream) розташована так звана лідерна послідовність, перед якою знаходяться гени білків Cas. Ці білкі мають нуклеазну активність. Фактично CRISPR є прокаріотичним аналогом системи інтерференції РНК (сайленсинга РНК) у еукаріот. Якщо спробувати сказати про роботу цієї системи дуже коротко, то воно виглядає так. Коли в бактерію потрапляє чужорідний генетичний матеріал, наприклад ДНК бактеріофага, фрагмент цієї ДНК копіюється і вставляється у CRISPR як новий спейсер. Спейсери складають імунологічну пам’ять бактерії чи археї, в якій зберігається інформація про минулі інфекції, і саме вона лежить в основі відповіді на повторне вторгнення подібних генетичних елементів. Після інтеграції в CRISPR частин чужорідних генетичних елементів потрібно перевести їх в форму, здатну націлювати білки Cas на послідовності-мішені для їх розпізнавання і руйнування. Такою формою служить напрямна crРНК (CRISPR-РНК), яка містить унікальну послідовність, комплементарну певній мішені. Спочатку низка повторів і спейсерів CRISPR транскрибується в єдиний довгий транскрипт – пре-crРНК, який далі розрізається на короткі crРНК. Більшість повторів в CRISPR є паліндромами, тому відповідні їм ділянки пре-crРНК формують шпильки. У багатьох випадках саме ці шпильки розпізнаються білками Cas, які перетворюють пре-crРНК на crРНК. Як правило, транскрипція CRISPR залежить від лидерної послідовності і відбувається постійно, але з низькою швидкістю. Однак швидкість значно збільшується в стресових умовах або при зіткненні клітини з фагами, забезпечуючи їй швидкий і ефективний захист. На стадії інтерференції crРНК зв’язуються зі своїми мішенями за рахунок спарювання комплементарних нуклеотидів і, таким чином, спрямовують ендонуклеази Cas на розрізання та руйнування мішені. Формування комплексу crРНК і білків Cas забезпечує руйнування комплементарних crРНК послідовностей нуклеїнових кислот. Хоча мішенями, в основному, є дволанцюгові ДНК, деякі системи CRISPR-Cas можуть руйнувати комплементарні одноланцюгові РНК. У спрощеному вигляді робота цієї системи зображена на першому малюнку. Таким чином і працює система CRISPR-Cas, і на її основі американські дослідники розробили систему PAC-MAN (Prophylactic Antiviral CRISPR in huMAN cells). Для цієї системи вони використали CRISPR-Cas13d систему з бактерії Ruminococcus flavefaciens. Cas13d це нещодавно виявлена РНК-ендонуклеаза. Також вони підібрали декілька crРНК для консервативних у багатьох коронавірусів і навіть вірусів грипу ділянок геномної РНК. Їх система успішно спрацювала проти РНК коронавірусів і вірусів грипу у культурі клітин епітелію легенів людини. Схематично це показано на другому малюнку. Але автори наприкінці роботи вірно зазначають, що є проблема – як доставити цю систему у клітини живої людини. Пропонується декілька стратегій, і якщо ця проблема буде вирішена, ми отримаємо дуже вдалу систему профілактики і лікування вірусних захворювань. Якщо когось цікавить текст цієї піонерської роботи, він тут.
Другий чудернацький противірусний препарат – це малі інтерферуючі РНК (міРНК), які можуть допомогти руйнувати РНК вірусів шляхом сайленсинга РНК. Сайленсинг РНК є функціональним еквівалентом системи CRISPR прокаріот, але він відбувається в еукаріотичних клітинах і працює він зовсім інакше. Сайленсинг, тобто «приглушення» РНК (RNA silencing), також відоме як пост-транскрипційне глушення генів або інтерференція РНК, індукується дволанцюговою РНК. В результаті цього процесу руйнується матрична РНК, яка має таку ж послідовність нуклеотидів, як і ініціаторна дволанцюгова РНК. Дволанцюгова РНК розпізнається комплексом білків, до складу якого входить фермент, РНКаза III, який зветься Dicer і є специфічним до дволанцюгової РНК, розрізаючи її на фрагменти довжиною 20–25 пар нуклеотидів (малі інтерферуючі РНК, міРНК). Фрагмент міРНК з’єднується з певними білками цитоплазми, формуючі комплекс RISC (RNA-induced silencing complex). У цьому комплексі дволанцюгова міРНК розплітається, і (–) ланцюг залишається в комплексі, що активує цей комплекс. Мінус-ланцюг РНК в комплексі націлюється на мРНК на ділянку, що має комплементарні нуклеотиди, і далі в цьому регіону мРНК руйнується. Схематично сайленсинг РНК зображено на третьому малюнку. Є велика спокуса знайти або синтезувати малі РНК, які будуть індукувати руйнуванні вірусних мРНК. Цікаво зазначити, що багато вірусів кодують власні міРНК, які спричиняють руйнування мРНК клітини-хазяїна, регулюючі її метаболізм у потрібний для реплікації вірусів бік. Щодо використання міРНК у якості агентів проти коронавірусів, я знайшов роботу турецьких авторів, у якій аналізується вплив міРНК з геному SARS-CoV-2 на гени людини. Передбачається, що в свою чергу міРНК геному людини можуть бути спрямовані на гени коронавірусу. Але поки що останній абзац статті звучить так: «Однак всі ці терапевтичні можливості потребують подальших механістичних оцінок, щоб зрозуміти, як вони регулюють взаємодію вірус-господар. Отже, для подальших досліджень in vitro, ex vivo та in vivo потрібно буде перевірити кандидатів серед міРНК для інфекції SARS-CoV-2». Так що все це в майбутньому. Ось текст статті.
Отакий от з SARS-CoV-2 нам клопіт, малята, любі хлопчики й дівчата. Не перейматися усім цим занадто і пам’ятайте, що гірше завжди попереду. Тоді і буде вам щастя.
Продовження фахових розповідей про збудник COVID-19 (а також його родичів та друзів) від автора університетського підручника “Вірусологія”, доцента нашої кафедри Сергія Шамрая (доступний за посиланням). Читайте також першу та другу частини на нашому сайті, якщо Ви досі цього не зробили.
Ось і третій есей щодо коронавірусів. Чи мабуть навіть невелика оповідь. Поговоримо про інтимне життя SARS-CoV і SARS-CoV-2. Тому, хто очікує, що мова буде про щось непристойне, нагадаю, що слово «інтимний» має багато відтінків, зокрема «неплатонічний». А взаємодію цих вірусів з клітинами нашого організму важко назвати платонічними. До того ж до вірусів неможливо застосувати слово «життя», принаймні у побутовому значенні.
Мова буде йти про деякі подробиці циклу реплікації двох вірусів. Вони загалом близькі за складом, використовують однакову стратегію паразитування у клітині хазяїна, але SARS-CoV вивчений набагато краще (його вивчають з 2002 року), тому багато відомостей ми обґрунтовано можемо екстраполювати на SARS-CoV-2.
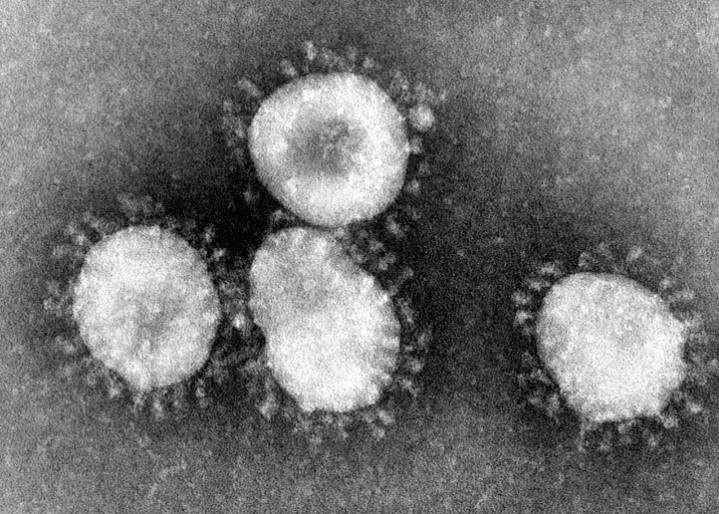
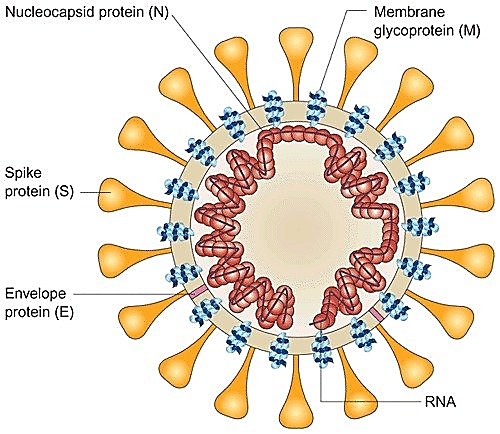
Нагадаю, вірусна частка (віріон) SARS-CoV-2 містить геном с позитивно сенсовою РНК (+РНК), яка укладена у спіраль разом з білком N, і цей нуклеокапсид оточений ліпопротеїновою мембраною (оболонкою), на який добре помітні вирости, які нагадують сонячну корону. Ці вирости інколи називають «шипами», але така назва не зовсім коректна, тому що вони не гострі і не жорсткі. Хоча англійська їх назва spike також означає шип, гвіздок, костиль (якім щось прикріплюють) тощо. Але я вважаю за краще звати їх виростами або виступами. Хоча можливо вирости також недобре, тому що вони не ростуть. Але хай вже буде. Нижче наведені електронна мікрофотографія віріонів і схематичне зображення їх структури.
(+)РНК-геном означає, що вірусна РНК є з одного боку геномом, а з іншого боку вона є матричною РНК (мРНК) і може безпосередньо транслюватися рибосомами з утворенням пептидних ланцюгів. Тобто безпосередньо на вірусному геномі синтезуються білки. Негативно сенсова РНК ((-)РНК) комплементарна до (+)РНК.
Дозволю собі ліричний відступ. Україно- і російськомовні біохіміки і молекулярні біологи часто чомусь полюбляють мРНК називати інформаційною РНК (іРНК), що є безглуздим. У англомовній літературі матричну РНК називають messenger RNA (mRNA), мессенджер-РНК, тобто РНК-посильний. Дійсно, вона є посильним між геномною ДНК і рибосомами, які синтезують на її матриці білок. Вона містить інформацію про послідовність амінокислотних залишків у поліпептидному ланцюгу. Але так само інформацію щодо послідовності амінокислот містить і (-)РНК, але у комплементарному вигляді! Тобто (-)РНК також є інформаційною, і багато вірусів мають саме геноми у вигляді (-)РНК! Але безпосередньо вона не може правити за матрицю для синтезу пептидів, і за визначенням не є інформаційною РНК. Саме тому називати мРНК інформаційною РНК є нонсенсом. Але багато наших шановних біохіміків і молекулярних біологів чомусь на це не зважають. Ну, загалом це не мій клопіт.
Коли коронавірусні частки потрапляють у потенційного хазяїна (наприклад, у когось з нас), перше, що їм треба зробити – це потрапити усередину сприйнятливих клітин. В першу чергу їм потрібно перетнути плазматичну мембрану, яка оточує кожну нашу клітину. А це – не тривіальна задача, тому що мембрана є напрочуд надійним бар’єром, який не дозволяє потрапляти до клітини тому, що туди потрапити не повинно. Ось тут віруси вдаються до хитрощів. Для приєднання до поверхні клітини вони використовують поверхневі білки-рецептори, які в нормі виконують важливі для клітини функції. І після зв’язування з рецептором вірус поглинається клітиною.
Вірус має так звані білки прикріплення, або антирецептори, які специфічно підходять до певного рецептора, як ключ до замка. Власне кажучи, віруси і здатні заражати тільки ті клітини, на поверхні яких є відповідні рецептори. І, наприклад, вірус імунодефіциту людини приєднується тільки до білків (CD4), які мають деякі клітини імунної системи, але не мають наприклад клітини легень. І навпаки, SARS-CoV і SARS-CoV-2 інфікують клітини легень, але не інфікують клітини імунної системи, тому що на останніх немає потрібного рецептора.
Рецептором, з яким зв’язується віріон SARS-CoV-2, є ангіотензинперетворюючий фермент 2 (angiotensin-converting enzyme 2, ACE2). Цей же рецептор використовує і збудник атипової пневмонії SARS-CoV. З боку вірусних часток білком прикріплення є як раз білок, який формує вирости на поверхні вірусних часток (білок S). Цікаво зазначити, що збудник близькосхідного респіраторного синдрому MERS-CoV у якості рецептора використовує іншій білок – DPP-4.
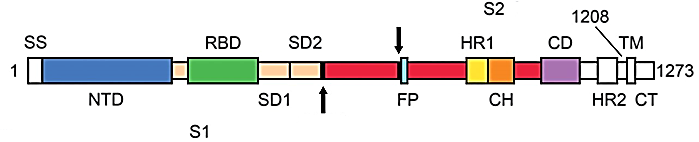
Білок S є тримером, тобто формується трьома однаковими поліпептидними ланцюгами. Він привертає багато уваги дослідників, тому що знаходиться на поверхні вірусної частки і можна очікувати, що саме з ним будуть взаємодіяти наївні В-лімфоцити, щоби перетворитися на клітини плазми крові і почати виробляти противірусні антитіла. Тобто саме білок S може бути антигеном при створенні вакцини проти SARS-CoV-2. Доменна структура цього білка наведена нижче.
Доменна структура білка S. S1, субодиниця зв’язування з рецептором; S2, субодиниця злиття мембран; SS, сигнальна послідовність; TM, трансмембранний домен; NTD, N-термінальний домен; FP, пептид злиття; HR1, гептадний повтор 1; HR2, гептадний повтор 2; RBD – домен зв’язування рецептора; SD1 і SD2, субдомени 1 і 2; CH, центральна спіраль; CT – цитоплазматичний хвіст. Стрілки вказують на сайти розрізання протеазою.
Кого цікавить детальна інформація щодо тривимірної структури білка S і її динаміки, вам сюди, сюди і сюди.
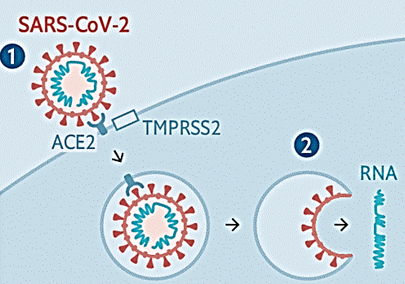
Коли вірус приєднується до рецептора, клітина вважає, що рецептор активувався природнім лігандом, і поглинає цей рецептор шляхом ендоцитозу з утворенням ендосоми. Але замість природного ліганду до клітини потрапляє вірусна частка! Тут важливо зрозуміти, що на цьому етапі вірусна частка потрапила в середину клітини, але не в цитозоль. Вірусна частка залишається оточеною мембраною ендосоми, а геном вірусу залишається оточеним ліпопротеїновою оболонкою власної вірусної частки. Щоби потрапити в цитоплазму, повинно відбутися злиття мембрани ендосоми з оболонкою вірусної частки, і саме тоді нуклеокапсид потрапить до цитозолю так би мовити автоматично.
Це відбувається наступним чином. Передбачається, що протеаза хазяїна TMPRSS2 розрізає рецептор ACE2, це потрібне для ендоцитозу і звільняє ділянку мембрани ендосоми. Надалі білок S також розрізається протеазою хазяїна (відбувається праймування (priming) білка S) між субодиницями S1 і S2. Як ми бачили на малюнку вище, цей білок має два потенційних сайти розрізання протеазою. Але у спеціалістів є поки що деяка непевність, які саме протеази розщеплюють білок S. Можлива участь тієї ж TMPRSS2, або це можуть бути фурін, трипсин, катепсин чи протеаза, схожа на трипсин в дихальних шляхах людини (human airway trypsin-like protease). Але в деяких публікаціях зовсім не вказують на те, що TMPRSS2 розрізає рецептор ACE2; ця протеаза тільки розрізає білок S у двох сайтах, активуючі злиття мембран.
Як би там не було, субодиниця зв’язування з рецептором S1 відрізається, активується субодиниця злиття мембран S2 і відбувається злиття мембран оболонки віріона і ендосоми. І геном вірусу нарешті опиняється у цитоплазмі, як показано на малюнку:
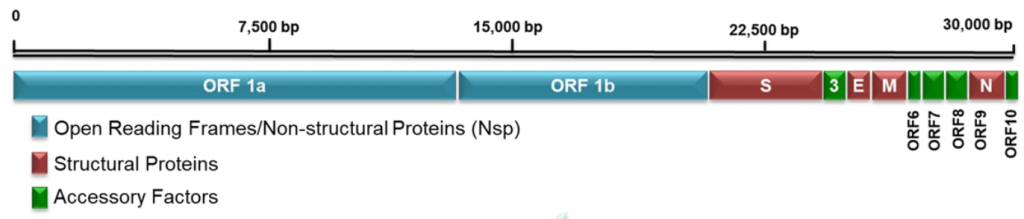
Схематичне зображення генома SARS-CoV-2 наведено нижче на малюнку. Нагадую, ORF означає відкриту рамку зчитування (open read frame).
Оскільки геном вірусу є одночасно мРНК, зразу ж починається його трансляція, тобто рибосоми клітини хазяїна, натрапивши на нього, починають синтезувати вірусні білки. На наведеній схемі різним кольором показані неструктурні, структурні і допоміжні білки SARS-CoV-2.
Тут треба знову зробити ще один ліричний відступ. У вірусів з геномами РНК є проблема. Фермент, якій повинен реплікувати цю РНК і здійснювати транскрипцію (синтез мРНК на (-)РНК), є РНК-залежною РНК-полімеразою. Але наші клітини його не мають! Ну немає у нас синтезу РНК на матриці РНК. Неможна сказати, що живі організми зовсім не мають РНК-залежної РНК полімерази, її мають рослини і деякі протисти. Але віруси цей фермент використовувати не можуть. Вони повинні кодувати свій власний фермент. Деякі фахівці навіть пропанують для вірусного ферменту назву РНК-залежна РНК полімераза (RNA-dependent RNA polymerase), а для ферменту живих організмів назву РНК-спрямована РНК полімераза (RNA-directed RNA polymerase). Але зараз щодо вірусів використовують обидві назви, нажаль. Таким чином, вірусу цей фермент потрібен для транскрипції і трансляції, і віруси або мають його в своїх вірусних частках (віруси з (-)РНК-геномами і з геномами, представленими дволанцюговими РНК), або він зразу ж синтезується у клітині на геномі у вірусів з (+)РНК-геномами, такими як SARS-CoV-2.
Але у вірусів з (+)РНК-геномами є ще одна проблема. Їх геном, як ми знаємо, фактично є мРНК, але ця РНК поліцистронна, тобто має декілька відкритих рамок зчитування, кожна з яких має стартовий кодон і стоп-кодон. От стоп-кодон і є проблемою. У нашого коронавірусу зразу ж буде транслюватися перша відкрита рамка зчитування (ORF1a, див. малюнок). Але коли зустрінеться її стоп-кодон, рибосома від’єднається. А правіші (downstream) рамки зчитування, як же їх транслювати? Віруси вирішують цю проблему різними шляхами, дещо про це сказано у нашому з Дмитром Вікторовичем Леонтьєвим підручнику.
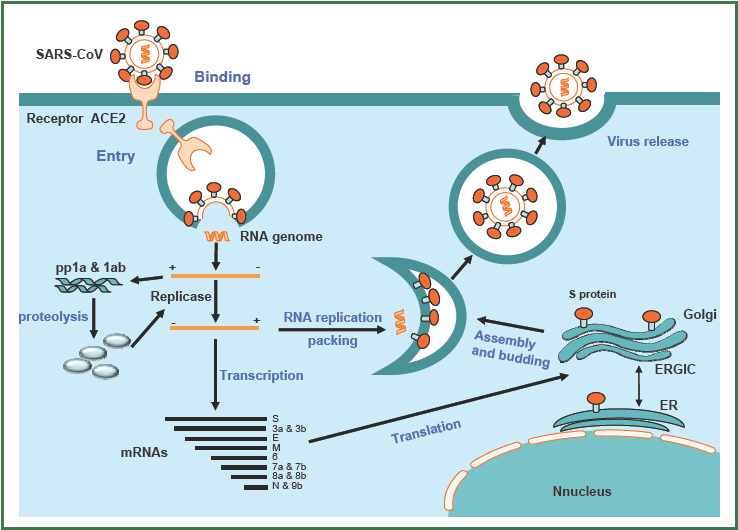
Що стосується SARS-CoV-2, то після попадання в клітину і «роздягання» геному відбувається кеп-залежна трансляція ORF1a з утворенням поліпептиду pp1a. Крім того, завдяки «слизькій» ділянці на РНК біля кінця ORF1a у 25-30% відбувається зсув рамки зчитування (-1 фреймшифтінг), завдяки чому транслюються ORF1a і ORF1b з утворенням більшого поліпептиду pp1ab. Надалі відбувається автокаталітичне розрізання цих поліпротеїнів з утворенням численних функціональних білків. Знаменно, що серед цих білків є РНК-залежна РНК полімераза, яка здійснює транскрипцію зокрема з утворенням субгеномних РНК (що є ще одним способом вирішення проблеми стоп-кодонів) і реплікацію генома. Схематично цикл реплікації SARS-коронавірусів зображений нижче.
Я не буду перераховувати усі численні вірусні білки, які синтезуються в клітині. Ці білки виконують численні функції, починаючі від запобігання імунної відповіді клітини і примушення клітини синтезувати вірусні білки, а не свої власні, і закінчуючи утворенням нових вірусних часток. Детальний аналіз білків з відомими і невідомими функціями наведено тут.
Коли у клітині буде напрацьована певна порогова кількість структурних вірусних білків і нових вірусних геномів, нові вірусні частки самозбираються і покидають клітину. Цікаво, що вірусні білки, які потраплять до оболонки вірусних часток, остаточно синтезуються і модифікуються в комплексі Гольджі. Новоутворені нуклеокапсиди брунькуються через мембрану бульбашок Гольджі, так що віріони придбавають власну мембрану, і оточені зверху ще однією мембраною. Надалі вони вивільняються з клітини шляхом екзоцитозу. Пишуть, що одна клітина може вивільнити 100-1000 вірусних часток, а потім гине.
А ми продовжуємо боротися з мракобіссям та невіглаством доступними для нас засобами. До Вашої уваги – другий есей доцента Сергія Шамрая про те, що всіх цікавить, але на чому мало хто розуміється. Першу частину читайте тут. Якщо ж Ви бажаєте набути комплексного уявлення про сучасну вірусологію – завантажуйте підручник Сергія Шамрая “Вірусологія” на нашому сайті!
Перший коронавірус у людини (HCoV-229E) був виявлений у 1965 році як збудник ГРЗ. Коронавіруси не привертали особливої уваги вірусологів і епідеміологів до 2002 року, до появи збудника атипової пневмонії SARS-CoV. Наразі відомо сім коронавірусів, які інфікують наш вид. Усі вони належать до роду бетакороновірус – одного з чотирьох родів підродини ортокоронавірусів у родині коронавірусів. До речі, вірусологи не надто заморочуються пошуком цікавих назв, і інші три роди звуться просто альфакоронавірус, дельтакоронавірус і гаммакоронавірус.
Усі сім коронавірусів, які вражають людей, мають походження від кажанів або гризунів, як показано на першому малюнку нижче.
Спалахи бетакоронавірусів у людини завжди включали попереднє потрапляння до тварин, інших ніж кажани. SARS-CoV та MERS-CoV передавались безпосередньо людям від вівер та верблюдів-дромадерів відповідно, SARS-CoV-2 ми отримали від панґолінів. На малюнку як HCoV зазначені різні коронавіруси людини (Human coronavirus), які повсюдно поширені і викликають сезонні застуди або більш-менш важкі ураження дихальних шляхів. На малюнку також показаний SADS‐CoV – вірус важкої діареї свиней (swine acute diarrhoea syndrome coronavirus), який у 2016 році викликав спалах фатальної хвороби свиней на фермах у Китаї, провінції Guangdong. Про те, що цей вірус інфікував людей, я ніякої інформації не знайшов.
Треба зазначити, що нам є чим пишатися. Коронавіруси – це на якесь там абищо. Вони мають найбільший геном серед усіх РНК-геномних вірусів, до 33 тисяч нуклеотидів одноланцюгової позитивно смислової РНК ((+)РНК). А наш улюблений SARS-CoV-2 має геном трішки менше ніж 30 тисяч нуклеотидів. Вірогідно, близько 33 тисяч нуклеотидів є граничним розміром РНК-геномів, які дозволяє матінка-природа для біологічних об’єктів (ДНК-геноми вірусів можуть мати до 1,9 млн пар нуклеотидів). Більші РНК-геноми мабуть будуть занадто нестабільними. Я не маю сумніву, що кожен біолог знає, чому геном РНК не може бути дуже великим, і чому віруси з РНК-геномами еволюціонують набагато швидше, аніж віруси з ДНК-геномами.
Цікавим питанням, яке стосується власне SARS-CoV-2, є таке: чи інфікує він домашніх тварин, і якщо інфікує, то яких саме. Наразі відомо, що цей вірус неясним шляхом може інфікувати котів і тхорів, але майже не заражає собак, свиней, курей та качок. Мені не трапилися експериментальні дані щодо хом’яків і морських свинок. Чому саме так, достеменно невідомо, але у зоопарку Нью-Йорка доглядач примудрився заразити навіть тигра.
Загалом для котів давно відомі коронавірусні хвороби, інколи з дуже важкими наслідками. Але котів заражають альфакоронавіруси, а нас, як ми знаємо, бетакоронавіруси. Яким чином бетакоронавірус SARS-CoV-2 заражає котів, повторюся, залишається загадкою. Якщо комусь цікаво прочитати оригінальну статтю про зараження тварин, ось вона.
І остання цікавинка. Спеціалістам не дає спокою різниця в кількості летальних наслідків зараження SARS-CoV-2 у різних країнах. В одному з досліджень були проаналізовані послідовності генів людини, які кодують рецептор АСЕ2. Нагадаю, що саме до цього рецептора прикріплюється білок виростів віріонів вірусу (білок S) для подальшого потрапляння в клітину людини. Додам маленьку подробицю, бо не все так просто. Після прикріплення, ще до потрапляння вірусної частки в клітину, рецептор АСЕ2 розрізається сериновою протеазою клітини хазяїна TMPRSS2, яка також розташована у мембрані. Це розрізання активує білок S таким чином, що він ініціює злиття ліпопротеїнової оболонки вірусу і мембрани клітини хазяїна, що і дозволяє нуклеокапсиду вірусу потрапити в цитозоль. І прикріплення, і розрізання критично важливі для успішного інфікування клітини. Так от, аналіз 7000 зразків генів, що кодують АСЕ2, у італійців виявив потенційні варіанти, які впливають на зв’язування, розщеплення і стабілізацію білків. Таким чином, генетичний фон може впливати на спостережувану клінічну мінливість. Також можливо оцінювати персональний ризик інфікування, що відкриває шлях до персоналізованих профілактичних заходів та терапевтичних варіантів. Кому цікава ця робота, вона ось тут.
У часи, коли інформаційний простір заповнено по самі вінця фейками, додумуваннями, хворобливими фантазіями “експертів з вірусології та епідеміології” (котрі ще вчора були експертами у політиці, міжнародних взаєминах та макроекономіці), суспільство потребує голосу здорового глузду. До Вашої уваги – текст автора найсвіжішого в Україні університетського підручника з вірусології, доцента нашої кафедри Сергія Шамрая.
У режимі самоізоляції-самоконсервації-самоліквідації зробив таке невеличке загально вірусологічне «есе» про коронавірус.
Спочатку хотів пісню написати, але нот не знаю. Тому пропоную це, можливо комусь цікаво. Переглядати тільки у віці після 18 років біологам, які хоч трохи пам’ятають вірусологію.
Let’s start. Коронавірус SARS-CoV-2 (від англ. Severe acute respiratory syndrome coronavirus 2), спочатку відомий як 2019-nCoV (novel coronavirus 2019) належить до родини коронавірусів (Coronaviridae), підродини ортокоронавірусів (Orthocoronavirinae), роду бетакоронавірусів (Betacoronavirus), підроду сарбековірус (Sarbecovirus). Ці віруси мають геном з одноланцюговою позитивно-сенсовою РНК, віріони вкриті ліпідною оболонкою, плеоморфні і мають характерні вирости на оболонці. Нижче наведені зображення цього зловмисника з камер спостереження і портрет, намальований криміналістами зі слів очевидців.
вірусні частки
реконструкція будови вірусу
Походження і еволюція. Декілька команд швидко встановили генетичну схожість SARS-CoV-2 і бетакоронавірусів підроду Sarbecovirus, який зустрічається на кажанах. Ідентичність послідовності SARS-CoV-2 і пов’язаного з SARS коронавірусу кажанів (SARSr-CoV; RaTG13) з провінції Юньнань, Китай, складає 96,2%. Цікаво, що новий вірус має не дуже велику подібність з коронавірусом атипової пневмонії (пам’ятаєте про такий?) SARS-CoV (близько 79%) і коронавірусом близькосхідного респіраторного синдрому MERS-CoV (близько 50%). Крім цього, критична ділянка гену, якій кодує білок виростів на поверхні віріонів, ідентична відповідній ділянці геному вірусів, виділених з панголінів. Але коли і як саме відбувся перехід вірусу кажан-панголін-людина, залишається невідомим.
Хоча була виявлена лише 4% варіабельність геномних нуклеотидів між SARS-CoV-2 та RaTG13, варіабельність з урахуванням нейтральних замін нуклеотидів складає 17%, що дозволяє припустити, що розбіжність між двома вірусами значно більша ніж раніше оцінювали. Виникнення нових варіацій функціональних ділянок у рецептор-зв’язуючому домені (RBD) виростів, що спостерігається у SARS-CoV-2 та вірусів панголіну SARSr-CoV, ймовірно, викликана мутаціями та природним відбором, крім рекомбінації. Популяційний генетичний аналіз 103 геномів SARS-CoV-2 показав, що ці віруси еволюціонували у два основні типи в залежності від амінокислоти, закодованої в певній ділянці геномів. Ці типи позначені як L (закодований лейцин) та S (закодований серин). Незважаючи на те, що тип L (∼70%) є більш поширеним, ніж тип S (∼30%), S тип виявився попереднім варіантом. Оскільки тип L був більш поширеним на ранніх стадіях спалаху в Ухані, частота типу L зменшувалася після початку січня 2020 року. Втручання людини може чинити більш сильний селективний тиск на тип L, який може бути більш агресивним і поширюватися швидше. З іншого боку, тип S, який еволюційно старіший і менш агресивний, міг би збільшити відносну частоту через відносно слабший селективний тиск.
Реплікація. Рецептором, з яким зв’язується віріон SARS-CoV-2, є ангіотензинперетворюючий фермент 2 (angiotensin-converting enzyme 2, ACE2). Можливо, є додаткові рецептори/корецептори. Цікаво, що цей же рецептор використовує і збудник атипової пневмонії SARS-CoV. Білкові вирости на віріонах обох вірусів мають дуже великий афінітет до ACE2. Надалі відбувається опосередкований рецептором ендоцитоз, злиття мембрани ендосоми і оболонки віріону і всі події, які трапляються з багатьма іншими (+)РНК-геномними вірусами, дивіться на малюнок нижче.Ключовим питанням є те, чому саме легені виявляються найбільш вразливим цільовим органом для вірусів. Однією з причин є те, що велика поверхня легені робить їх дуже чутливими до вірусів, що вдихаються, але є також біологічний фактор. Було продемонстровано, що 83% клітин, що експресують ACE2, є клітинами альвеолярного епітелію II типу (alveolar epithelial type II cells, AECII), що дозволяє припустити, що ці клітини можуть служити воротами для вірусної інвазії. Крім того, показано, що AECII, що експресують АСЕ2, мають високий рівень множинних процесів, які потрібні вірусу для реплікації. Це дозволяє припустити, що саме AECII, що експресують АСЕ2, полегшують реплікацію коронавірусу в легенях.
схема взаємодії вірусу з клітиною
Експресія рецептора АСЕ2 також спостерігається у багатьох не легеневих тканинах, включаючи серце, нирки, ендотелій та кишковик. Важливо, що АСЕ2 сильно експресується на поверхні епітеліальних клітин кишечника, функціонуючи як ко-рецептор для поглинання поживних речовин, зокрема для резорбції амінокислот з їжею. Тому прогнозується, що кишечник також може бути основним місцем потрапляння в організм SARS-CoV-2 і що зараження могло бути ініційовано вживанням їжі з ринку Ухань, передбачуваним місцем спалаху. Чи дійсно SARS-CoV-2 може заразити епітелій кишечника людини, має важливе значення для фекально-оральної передачі та стримування поширення вірусу. Слід зазначити, що згідно з деякими нещодавніми публікаціями, чи людина може отримати SARS-CoV-2, торкаючись поверхонь або предметів, що мають на собі вірус, а потім торкаючись власних слизових оболонок, ще не підтверджено.